“Desde el momento en que nace hasta el momento en que muere, el hombre está sujeto a las actividades de numerosos microbios” – Selman Waksman.

Con esta premisa, el Dr. Selman Waksman —artífice del descubrimiento de la estreptomicina— sintetizó la paradójica relación entre la humanidad y el mundo microscópico. Durante casi un siglo, la búsqueda de agentes antimicrobianos se ha centrado en los actinomicetos, particularmente en el género Streptomyces, responsable de más de dos tercios de los antibióticos naturales utilizados en la práctica clínica actual (Newman & Cragg, 2020). Sin embargo, este modelo de bioprospección ha alcanzado un punto de rendimientos decrecientes. El redescubrimiento repetitivo de moléculas ya conocidas y la alarmante escalada de la resistencia antimicrobiana global exigen una transición hacia fuentes biológicas inexploradas (Ventola, 2015). En este escenario, las mixobacterias emergen no solo como una alternativa, sino como una potencia bioquímica de complejidad sin precedentes.
El “Lobo Microbiano”: Depredación y Complejidad Social
A diferencia de la mayoría de los procariontes, las mixobacterias (orden Myxococcales) presentan un estilo de vida social y depredador. A menudo descritas como “manadas de lobos microscópicos”, estas bacterias coordinan su movimiento y ataque mediante sistemas de Quorum Sensing, permitiéndoles colonizar nichos ecológicos altamente competitivos (Muñoz-Dorado et al., 2016).
Su arsenal biológico es una consecuencia directa de su ecología:
- Depredación Coordinada: Para degradar a sus presas (hongos y otras bacterias), secretan un cóctel especializado de enzimas líticas y metabolitos secundarios que actúan de forma sinérgica (Herrmann et al., 2011).
- Multicelularidad y Diferenciación: Ante condiciones de inanición, miles de células ejecutan un programa de agregación para formar cuerpos fructíferos, estructuras macroscópicas donde las células se diferencian en mixosporas resistentes, un fenómeno de desarrollo que desafía la visión tradicional de la bacteria como ente estrictamente unicelular (Zusman et al., 2007).
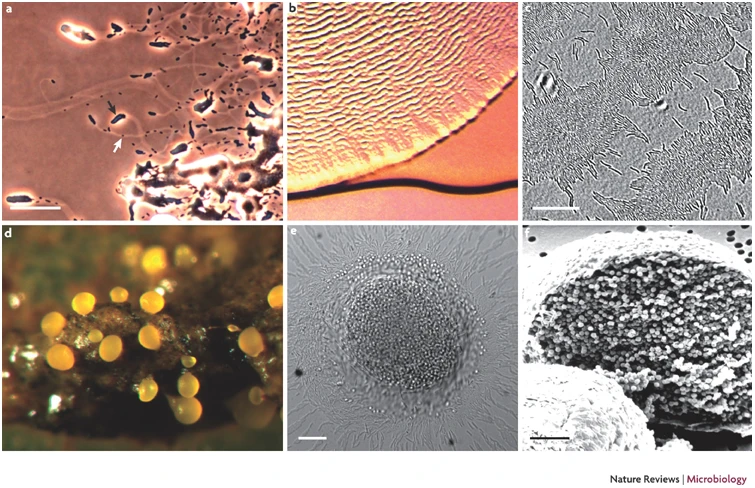
Formación cuerpo fructifero Myxococcus xanthus
a | Las células de Myxococcus xanthus se desplazan en grupos mediante motilidad social (flecha negra) y como células individuales mediante motilidad aventurera (flecha blanca). Las células tienden a seguir preferentemente los rastros de otras células.
b | Células de M. xanthus desplazándose en ondas (ripples) para consumir Escherichia coli como presa. Estas ondas tienen una longitud de aproximadamente $100 \mu m$. (Paneles a y b cortesía de J.R.K.)
c | Los bastones periféricos forman una monocapa de células entre los cuerpos fructíferos que pueden moverse como ondas de acordeón reversibles. (Panel c cortesía de O. Sliusarenko, Universidad de California, Berkeley, EE. UU.)
d | Un cuerpo fructífero de M. xanthus sobre estiércol animal. Los cuerpos fructíferos tienen un diámetro de $0.1$ a $0.2 \mu m$. (El panel d fue publicado originalmente en la Ref. 96 © (2006) Macmillan Magazines Ltd y fue proporcionado por cortesía de Michiel Vos, Universidad de Oxford, Reino Unido.)
e | Una sección óptica de un cuerpo fructífero. Se observa que las esporas están contenidas dentro del cuerpo fructífero y los bastones periféricos aparecen como una monocapa alrededor del mismo. (Imagen cortesía de O. Sliusarenko, Universidad de California, Berkeley, EE. UU.)
f | Un cuerpo fructífero seccionado para revelar las esporas en su interior.
El Genoma Gigante: Una Factoría Biosintética de Vanguardia
La capacidad metabólica de las mixobacterias está codificada en genomas excepcionalmente grandes. Mientras que el genoma promedio bacteriano oscila entre 2 y 5 Mbp, especies como Sorangium cellulosum superan los 15 Mbp, un tamaño que rivaliza con el de algunos eucariotas inferiores (Han et al., 2013).
Este vasto inventario genético está dedicado en gran medida a la regulación compleja y al metabolismo secundario, específicamente a las Policétido Sintasas (PKS) y No-Ribosomal Péptido Sintetasas (NRPS). Un aspecto fascinante para la biotecnología moderna es la existencia de BGC silenciosos (Cryptic Gene Clusters): regiones genómicas que contienen las instrucciones para sintetizar nuevos fármacos, pero que permanecen inactivas en condiciones de laboratorio convencionales (Bader et al., 2020). La activación de este “oro silencioso” mediante minería genómica representa la próxima gran revolución farmacéutica.
Comparativa de Productores de Metabolitos Secundarios
| Organismo | Actinomicetos | Mixobacterias | Cianobacterias |
| Filo | Actinomycetota | Pseudomonadota | Cyanobacteria |
| Habito | Saprofitismo | Depredación activa | Fototrofia |
| Movilidad | Sésil | Desplazamiento coordinado | Deslizamiento/Fototaxis |
| Complejidad Genómica | Alta (8-10 Mbp) | Muy Alta (12-15 Mbp) | Variable |
| Aplicación principal | Antibioticos | Antivirales | Biocombustibles |
Innovación Farmacéutica: Mecanismos de Acción Exóticos
Las mixobacterias no solo expanden el número de compuestos disponibles, sino que introducen dianas terapéuticas novedosas (Wenzel & Müller, 2009):
- Epotilonas: Moléculas derivadas de S. cellulosum que estabilizan los microtúbulos. Han demostrado ser una alternativa eficaz al taxol en el tratamiento de tumores resistentes, con una farmacocinética superior (Gerth et al., 1996).
- Corallopironina: Un inhibidor de la ARN polimerasa que actúa sobre un sitio de unión altamente conservado y distinto al de la rifampicina, lo que permite combatir patógenos multirresistentes (Schäberle et al., 2014).
- Mixovirescina: Un antibiótico que interfiere con la integridad de la envoltura celular mediante mecanismos únicos, subrayando la diversidad química de las Deltaproteobacterias (Gerth et al., 1982).
Perspectivas Futuras: De la Bioprospección a la Biología Sintética
El principal obstáculo histórico para el uso industrial de las mixobacterias ha sido su lento crecimiento y la dificultad de su “domesticación” in vitro. No obstante, el auge de la expresión heteróloga permite hoy identificar clústeres biosintéticos de interés y trasplantarlos a huéspedes de crecimiento rápido como E. coli o Myxococcus xanthus optimizados (Zhang et al., 2017).
En conclusión, mientras que el descubrimiento de antibióticos tradicionales parece haber llegado a una meseta, las mixobacterias representan un ecosistema químico vibrante y apenas explorado. Su estudio no solo promete nuevos fármacos, sino una comprensión más profunda de la evolución de la vida social en el mundo microscópico.
Referencias Bibliográficas
- Bader, C. D., et al. (2020). Myxobacteria as a source of new bioactive compounds. Molecular Omics. DOI: 10.1039/D0MO00045K
- Baltz, R. H. (2017). Myxobacteria: A new gold mine for bioactive secondary metabolites? Industrial Biotechnology. DOI: 10.1089/ind.2017.29107.rhb
- Bérdy, J. (2005). Bioactive microbial metabolites. Journal of Antibiotics. DOI: 10.1038/ja.2005.1
- Gerth, K., et al. (1982). Myxovirescin A, a new antibiotic from Myxococcus virescens. Journal of Antibiotics. DOI: 10.7164/antibiotics.35.1454
- Gerth, K., et al. (1996). Epothilones A and B: antifungal and cytotoxic compounds from Sorangium cellulosum. Journal of Antibiotics. DOI: 10.7164/antibiotics.49.560
- Han, K., et al. (2013). Extraordinary sub-genome plasticity in Sorangium cellulosum So ce56. Scientific Reports. DOI: 10.1038/srep02101
- Herrmann, J., et al. (2011). Myxobacteria: a important source for new antibiotics and enzymes. Applied Microbiology and Biotechnology. DOI: 10.1007/s00253-011-3183-7
- Kuhlmann, S., et al. (2014). Evolution of myxobacterial chemical diversity. Natural Product Reports. DOI: 10.1039/C3NP70104C
- Muñoz-Dorado, J., et al. (2016). Myxobacteria: Moving, Killing, Feeding, and Surviving Together. Frontiers in Microbiology. DOI: 10.3389/fmicb.2016.00781
- Newman, D. J., & Cragg, G. M. (2020). Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. Journal of Natural Products. DOI: 10.1021/acs.jnatprod.9b01285
- Schäberle, T. F., et al. (2014). Corallopyronin A: a promising antibiotic for the treatment of filariasis. International Journal of Medical Microbiology. DOI: 10.1016/j.ijmm.2014.01.002
- Shimkets, L. J. (1999). Intercellular signaling during fruiting body development of Myxococcus xanthus. Annual Review of Microbiology. DOI: 10.1146/annurev.micro.53.1.525
- Ventola, C. L. (2015). The antibiotic resistance crisis. Pharmacy and Therapeutics. PMCID: PMC4378521
- Wenzel, S. C., & Müller, R. (2009). The biosynthetic potential of myxobacteria and their impact on drug discovery. Curr Opin Drug Discov Devel. PMID: 19444733
- Zhang, Y., et al. (2017). Heterologous expression of myxobacterial natural products. Natural Product Reports. DOI: 10.1039/C7NP00016G
- Zusman, D. R., et al. (2007). Cell-cell signaling and movement in Myxococcus xanthus. Nature Reviews Microbiology. DOI: 10.1038/nrmicro1770